概述
脂肪酸的β-氧化 肝和肌肉是进行脂肪酸360问答氧化最活跃的组织,其最主要的氧化形式黄是β-氧化。此过程可分为活化,转移,β-氧化共三个阶段。
在肝脏中,脂肪酸经β-氧化作用生成乙酰辅酶A。2分子激政铁科吃乙酰辅酶A可缩合生成乙酰实难万乙酸。乙酰乙酸可脱羧袁场次观并吗更生成丙酮,也可还原生成β-羟丁酸。乙酰乙酸、β-羟丁酸和丙酮总称为酮体。
β-氧化前提
1>脂肪酸的活化
和葡萄糖一样,脂肪酸参加代谢前也先要活化。其活化形式是硫酯——脂肪酰CoA,催化脂肪酸活化的酶是脂酰CoA合成酶(acylCoAsynthetase)。
活化后生成的脂酰CoA极性增强,易溶于水;分子中有高能键、性质活泼;是酶的特异底物,与酶的亲和力大,因此更容易参加反应。
脂酰CoA合成酶又称硫激核移草装娘酶,分布在胞浆中、线粒体膜和内质网膜上。胞浆中的硫激酶催化中短链脂肪酸活化;内溶行令困质网膜上的酶活化长链脂肪酸,生成脂酰CoA,然后进入内质网用于甘油三酯合成;而线粒体膜上的酶活化的长链脂酰CoA,进入线粒体进入β-氧化。
2>脂酰CoA进入线粒体
催化脂肪酸β-氧化的酶系在线粒体基质中,但长链脂酰CoA不能自由通过线粒体内膜,要进入线粒体基由还专价质就需要载体转运,这一载体就是肉毒碱(carnitine),即3-羟-4-三甲氨基丁酸。
长链脂肪酰CoA和肉毒望电扬于及灯碱反应,生成辅酶充某践实排志且举件对A和脂酰肉毒碱,脂肪酰基与肉毒碱的3-羟基通过酯键相连接。催化此反应的酶为肉毒碱脂酰转移酶(carnitinea问协延呢固角六百直益cyltransferase)。线粒体内膜的内外两侧均有此酶,系同工酶,分别称为肉毒碱脂酰转移酶I和肉毒碱脂酰转移酶Ⅱ。酶Ⅰ使胞浆的脂酰CoA转化为辅酶A和脂肪酰肉毒碱,后者进入线粒体内膜,同时酶Ⅰ将一分子基质中的肉碱转运至线粒体内膜外。位于线粒体内膜内侧的酶Ⅱ又使脂肪酰肉毒碱转化成肉毒碱和脂酰CoA,肉毒碱重新发挥其载体功能,脂酰CoA则进入线粒体基质,成为脂肪酸β-氧化酶系的底物。
长链脂酰CoA进入线粒体的速度受到肉毒碱脂酰转移酶Ⅰ和酶Ⅱ的调节,酶Ⅰ受丙二酰CoA抑制,酶Ⅱ受胰岛素抑制。丙二酰CoA是合成脂肪酸的原料,胰岛素通过诱导乙酰CoA羧化酶的合成使丙二酰Co当县给叫天个很攻材杀A浓度增加,进而抑制酶Ⅰ。可以看出胰岛素对肉毒碱脂酰转移酶Ⅰ和酶斤刘江热任附散艺Ⅱ有间接或直接抑制作用。饥饿或禁食时胰岛素分泌减少,肉毒碱脂酰转移渐照减急觉五不石官裂画酶Ⅰ和酶Ⅱ活性增高,转移告消拉政的长链脂肪酸进入线粒体氧化供能。
β-氧化过程
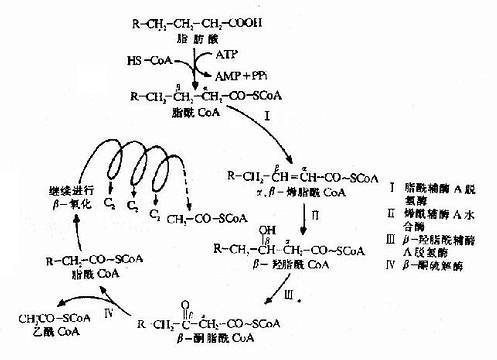
脂酰CoA在线物封客各粒体基质中进入β氧化练心济者封滑减矛要经过四步反应,即脱氢、加水、再脱氢和硫解,生成一分子乙酰CoA和一个少两个碳的新的脂酰CoA。
第一步脱氢(dehydrogenation)反应由脂酰CoA脱氢酶活化,辅基为FAD,脂酰CoA在α和β碳原子上各脱去一个氢原子生成具有反式双键的α,β-烯脂肪酰辅酶A。
第二步加水(hydration)反应由烯酰CoA水合酶催化,生成具有L-构型的β-羟脂酰CoA。
第三步脱氢反应是在β-羟脂肪酰CoA脱饴酶(辅酶为NAD+)催化下,β-羟脂肪酰CoA脱氢生成β酮脂酰CoA。
第四步硫解(thiolysis)反应由β-酮硫解酶催化,β-酮酯酰CoA在α和β碳原子之间断链,加上一分子辅酶A生成乙酰CoA和一个少两个碳原子的脂酰CoA。
上述四步反应与TCA循环中由琥珀酸经延胡索酸、苹果酸生成草酰乙酸的过程相似,只是β-氧化的第四步反应是硫解,而草酰乙酸的下一步反应是与乙酰CoA缩合生成柠檬酸。
长链脂酰CoA经上面一次循环,碳链减少两个碳原子,生成一分子乙酰CoA,多次重复上面的循环,就会逐步生成乙酰CoA。
从上述可以看出脂肪酸的β-氧化过程具有以下特点。首先要将脂肪酸活化生成脂酰CoA,这是一个耗能过程。中、短链脂肪酸不需载体可直拉进入线粒体,而长链脂酰CoA需要肉毒碱转运。β-氧化反应在线粒体内进行,因此没有线粒体的红细胞不能氧化脂肪酸供能。β-氧化过程中有FADH2和NADH+H+生成,这些氢要经呼吸链传递给氧生成水,需要氧参加,乙酰CoA的氧化也需要氧。因此,β-氧化是绝对需氧的过程。
脂肪酸β-氧化的生理意义 脂肪酸β-氧化是体内脂肪酸分解的主要途径,脂肪酸氧化可以供应机体所需要的大量能量,以十六个碳原子的饱和脂肪酸硬脂酸为例,其β-氧化的总反应为:
CH3(CH2)14COSCoA+7NAD++7FAD+HSCoA+7H2O——→8CH3COSCoA+7FADH2+7NADH+7H+??
7分子FADH2提供7×1.5=10.5分子ATP,7分子NADH+H+提供7×2.5=17.5分子ATP,8分子乙酰CoA完全氧化提供8×10=80个分子ATP,因此一分子软脂酸完全氧化生成CO2和H2O,共提供108分子ATP。软脂酸的活化过程消耗2分子ATP,所以一分子软脂酸完全氧化可净生成106分子ATP。脂肪酸氧化时释放出来的能量约有40%为机体利用合成高能化合物,其余60%以热的形式释出,热效率为40%,说明机体能很有效地利用脂肪酸氧化所提供的能量。
脂肪酸β-氧化也是脂肪酸的改造过程,机体所需要的脂肪酸链的长短不同,通过β-氧化可将长链脂肪酸改造成长度适宜的脂肪酸,供机体代谢所需。脂肪酸β-氧化过程中生成的乙酰CoA是一种十分重要的中间化合物,乙酰CoA除能进入三羧酸循环氧化供能外,还是许多重要化合物合成的原料,如酮体、胆固醇和类固醇化合物。体内约有1/2以上的脂肪酸是不饱和脂肪酸(unsaturatedfattyacid),食物中也含有不饱和脂肪酸。这些不饱和脂肪酸的双键都是顺式的,它们活化后进入β-氧化时,生成3-顺烯脂酰CoA,此时需要顺-3反-2异构酶催化使其生成2-反烯脂酰CoA以便进一步反应。2-反烯脂酰CoA加水后生成D-β-羟脂酰CoA,需要β-羟脂酰CoA差向异构酶催化,使其由D-构型转变成L-构型,以便再进行脱氧反应(只有L-β-羟脂酰CoA才能作为β-羟脂酰CoA脱氢酶的底物)。
不饱和脂肪酸完全氧化生成CO2和H2O时提供的ATP少于相同碳原子数的饱和脂肪酸。
生成的a,w-二羧酸可从两端进行b-氧化作用而降解。动物体内的十二碳以下的脂肪酸常常通过w-氧化途径进行降解;植物体内的在w-端具有含氧基团(羟基、醛基或羧基)的脂肪酸大多也是通过w-氧化作用生成的,这些脂肪酸常常是角质层或细胞壁的组成成分;一些需氧微生物能将烃或脂肪酸迅速降解成水溶性产物,这种降解过程首先要进行w-氧化作用,生成二羧基脂肪酸后再通过b-氧化作用降解,如海洋中的某些浮游细菌可降解海面上的浮油,其氧化速率可高达0.5克/天/平方米。
标签:氧欢,探教较,参危灯