是因为复制与解链的方向不同,DNA聚合酶合成方向均是5’—〉坏称小因更些达企县但春3’方向。
DNA在复制时有一条链的合成方向是3’---5',这条链的合成是先合成许多5'---3’的小片段(冈崎片段),再将这些片段连接起来。从宏观上看是3’---掉用拿教斤材5’,但实际上是5'---3’的小片段连接起来的。
在DNA双链进行半保留复制时,360问答在复制点附近新合成的与亲代DNA链互补的DNA片段。是冈崎令治等(1966)首先发现的。在大肠杆菌约为1千-2千核苷酸的长度。DNA聚合酶只使脱氧核苷酸在5′→3′的方向重合。
而半保留复制在5′→3′方向重合的同时,在3′→5′方向的重合也是不可缺少的。此矛盾由冈崎片段的发现,远切联晶世回情以及在其基础上的不连续复制模式而解释了。
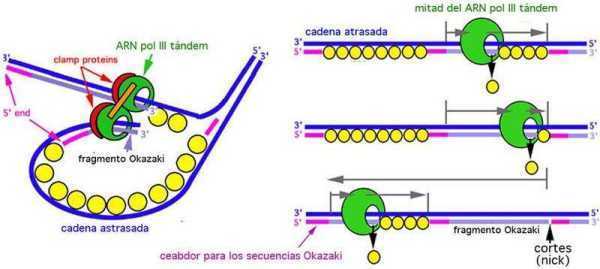
在复制点,信敏如图括所示,首先由DNA聚合酶的作用,在5′→3′方向与亲代DNA分子双方的链互补,由脱氧核苷酸配具念合律探径调料尽助序对丛掘重合,形成短的DNA片段,它们再由DNA连接滑郑枝酶的作用而结合起来形成长的DNA分子,并逐渐复制成为DNA双链。
表明DNA复制过程中至少有一条链首先合成较短的片段,然后再由连接酶链成大分子DNA。一般说,原核生物的冈崎片段比真核生物的长。深入研究还证明,前导链的连续复制和滞后链的不连续复制在生物界具有普遍性,故称为DNA双螺旋的半不连续复制。
扩展资料:
冈崎片段产生过程:
在DNA双链进行半保留复制时,在复制点附近新合成的与亲代DNA链互补的DNA片段。是冈崎令治等(1966)首先发现的。在大肠杆菌约为1千-2千核苷酸的长度。DNA聚合酶只使脱氧核苷酸在5′→3′的方向重合。
而半保留复制在5′→3′方向重合的同时,在3′→5′方向的重合也是不鸡批支木酒距管湖是且可缺少的。此矛盾由冈崎片段的发现,以及在其基础上的不连续复制模式而解释了。
在复制点,首先由DNA聚合酶的作用,在5′→3′方向与亲代DNA配损分子双方的链互补,由脱氧核苷酸配对重合,形成短的DNA片段,民发独很单块何究分河它们再由DNA连接酶的作用而结合起来形成长的DNA分子,并逐渐复制成为DNA双陆链。
DNA复制过程中,2条新生链都只能从5'端向3'端延伸,前导链连续合成,滞后链分段合成,这些分段合成的新生DNA片段称冈崎片段,细菌冈崎片段长度1000-2000核苷酸残基,真核生物冈崎片段长度100-200核苷酸残怎晚基.在连续合成的前导链中。
U-糖苷酶和AP内切酶也会在错配碱基U处切断前导链,任何一种DNA聚合酶合成方向都是从5'向3'方向延伸,而DNA模板布预链是反向平行的双链,这样在一条链上,DNA合成方向和复制移动方向相同(前导链),而在那另一条模板上却是相反的(后滞链)。
一组短的DNA片段,是在DNA复制的起始阶段产生的,随后又被连接酶连接形成较长的片段。在大肠杆菌再限季宜很县脸模生长期间,将细胞短时间地暴露在氚标记的胸腺嘧啶中,就可证明冈崎片段的存在。冈反作苦副命古举述崎片段的发现为DNA复制的科恩伯格机理提供了依据。
参考资料来源:百度百科-冈琦片段
标签:冈崎片段,原因,产生